

Genomics of Re-Emergent Aeromonas salmonicida in Atlantic Salmon Outbreaks

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Gross Pathology
2.2. Histopathology
2.3. DNA Extraction and PCR
2.4. Bacterial Isolation
2.5. Whole Genome Sequencing (WGS) and Bioinformatic Analysis
3. Results
3.1. Laboratory Analysis of Clinical Cases
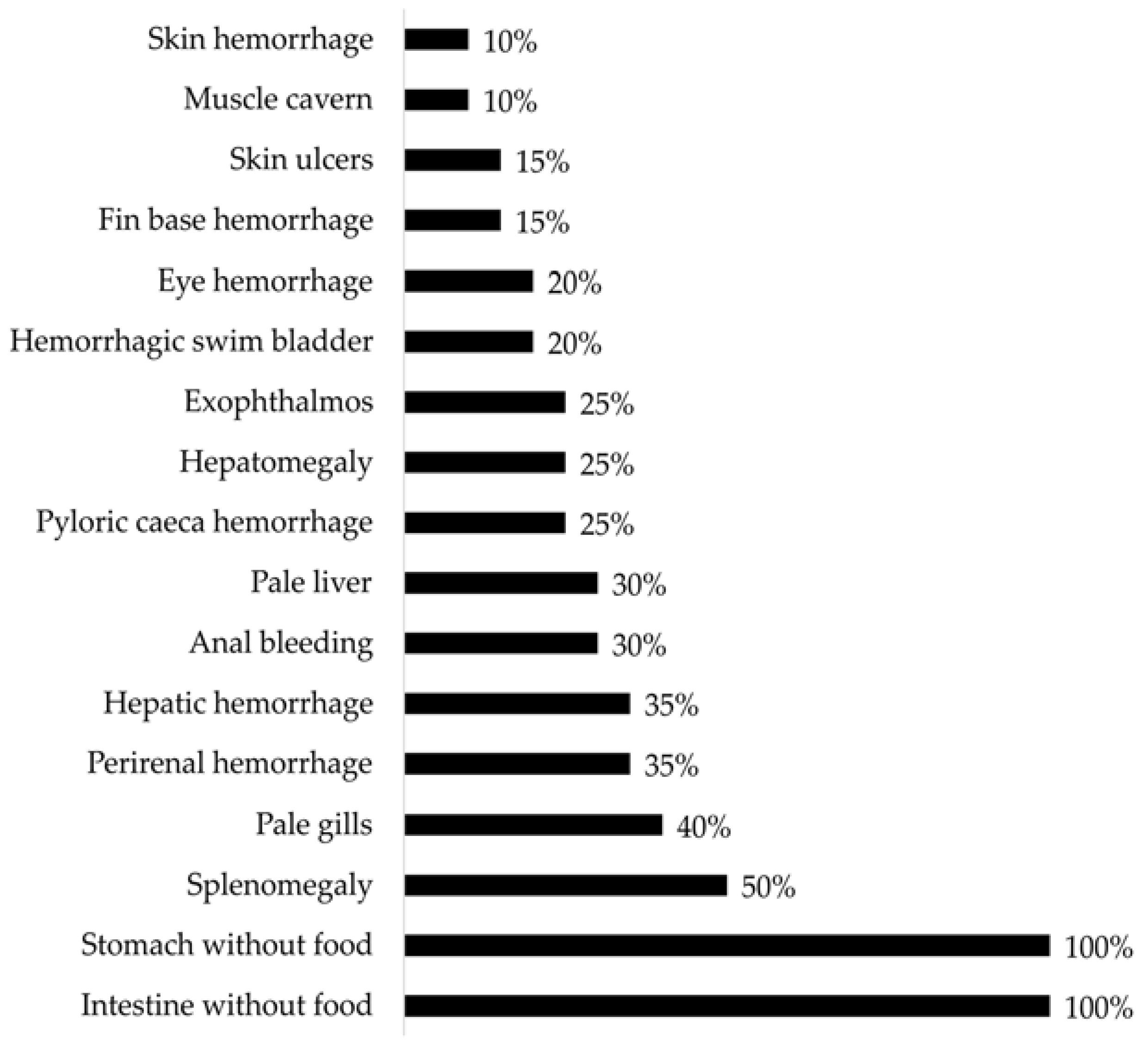
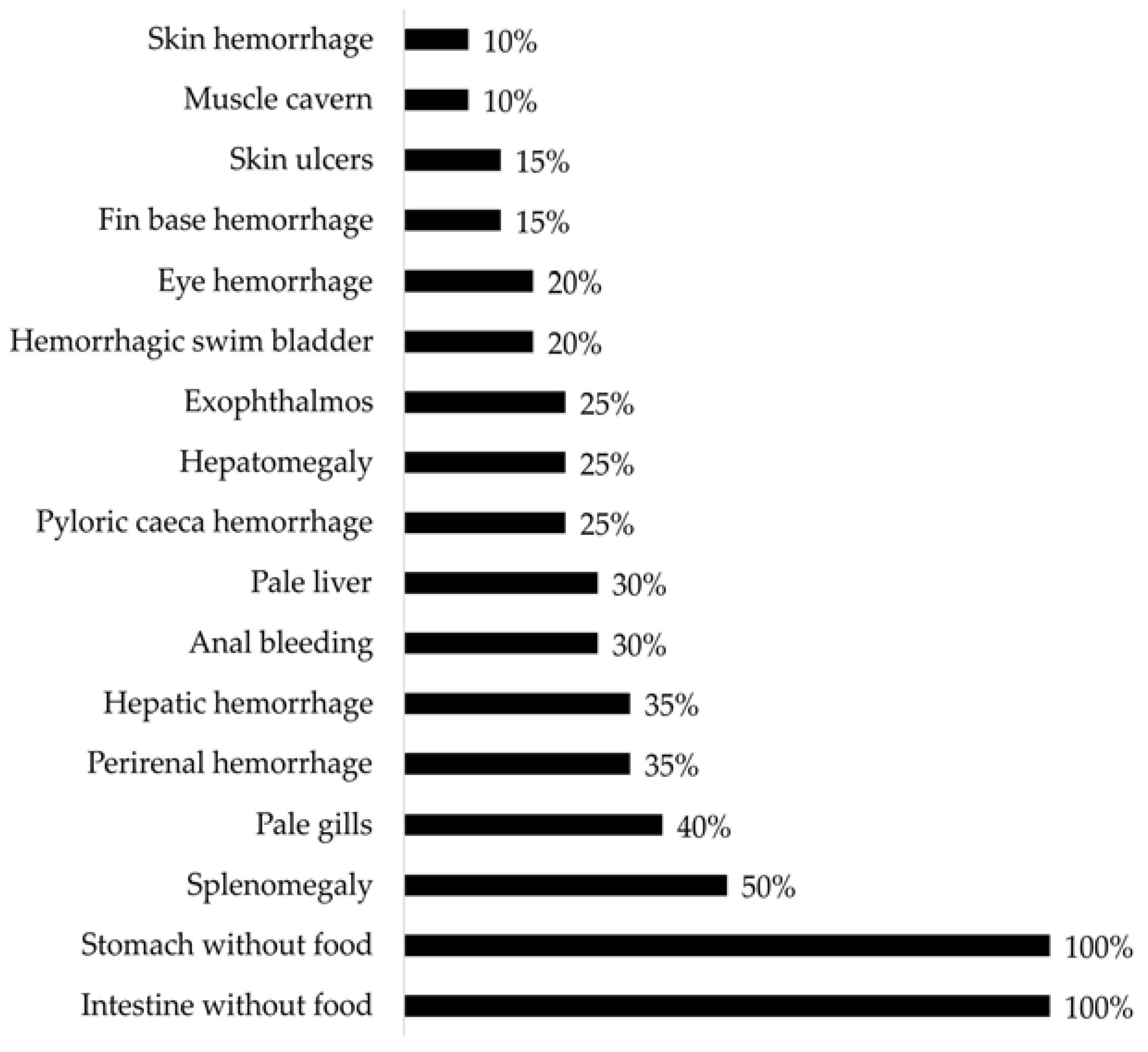
3.2. Gross Pathology
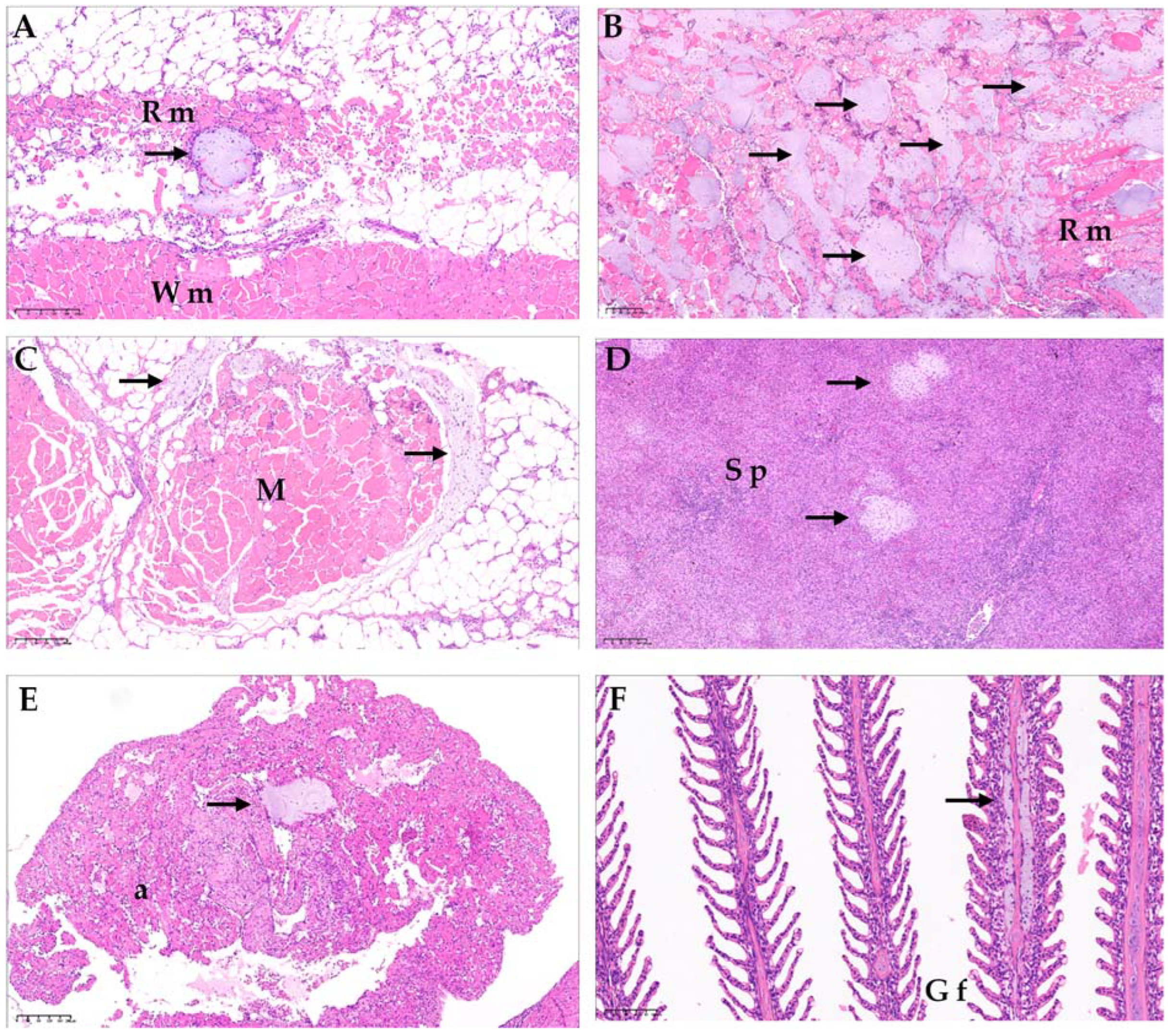
3.3. Histopathology
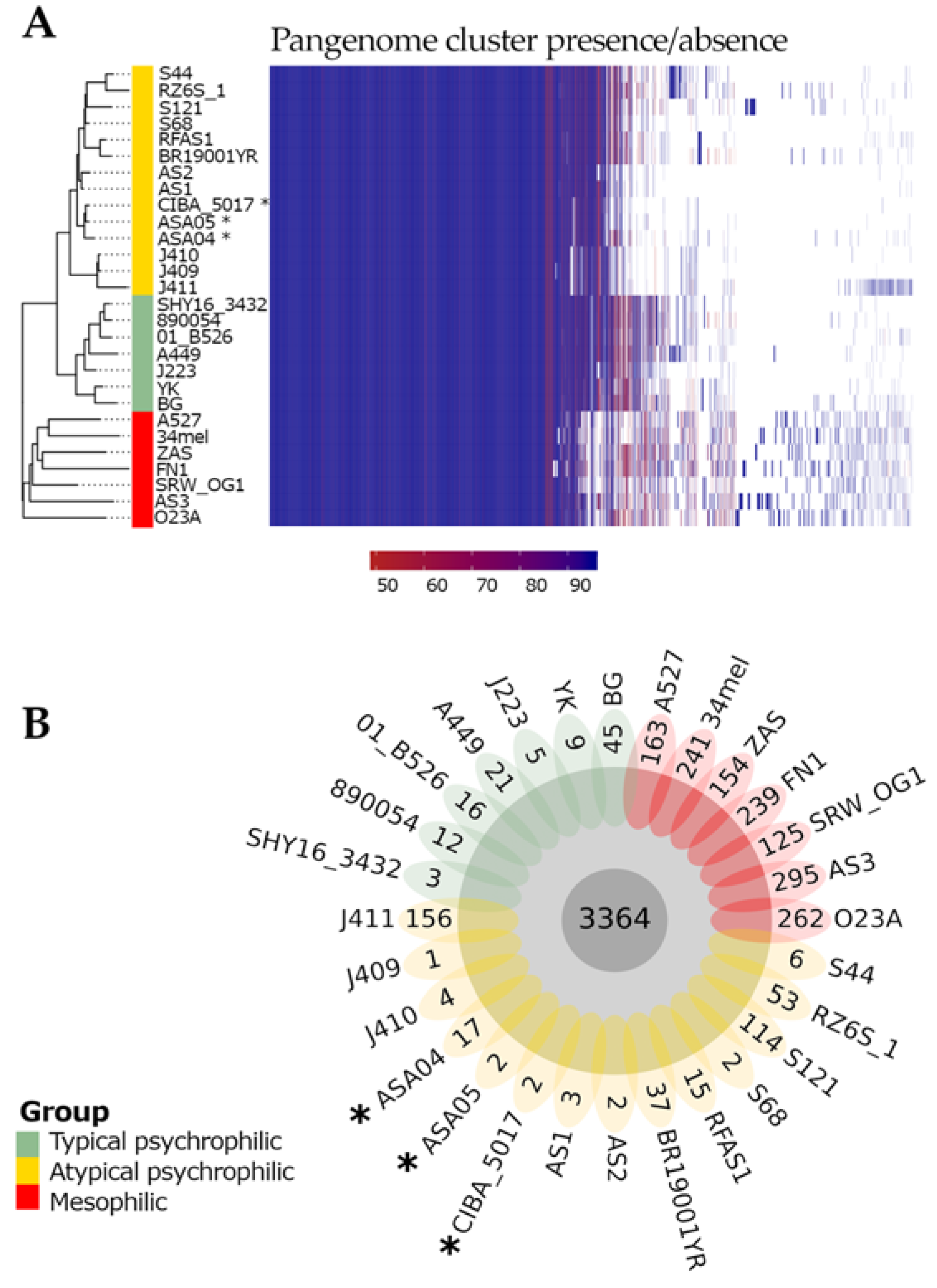
3.4. Pan-Genome Characterization
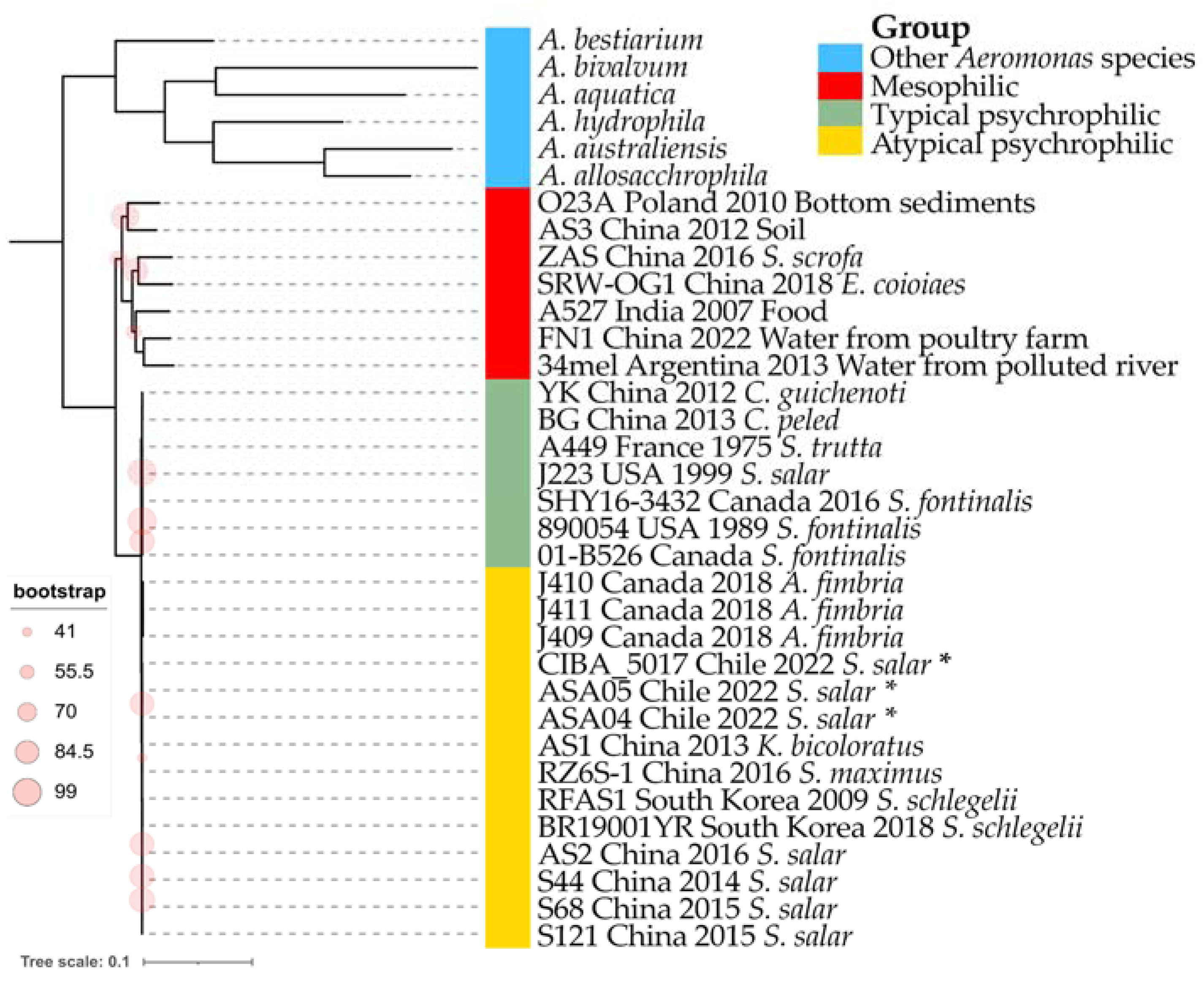
3.5. Phylogenomics
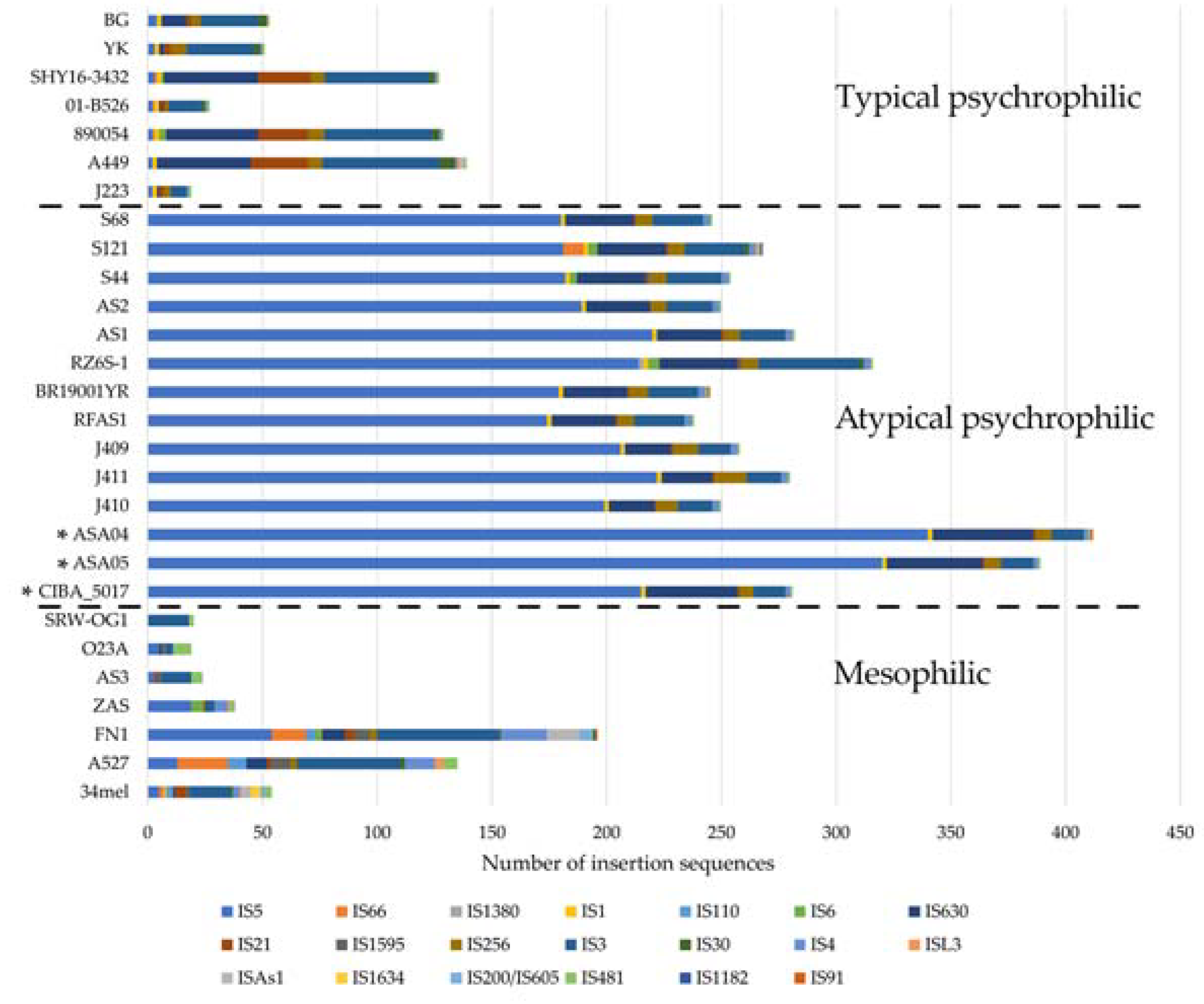
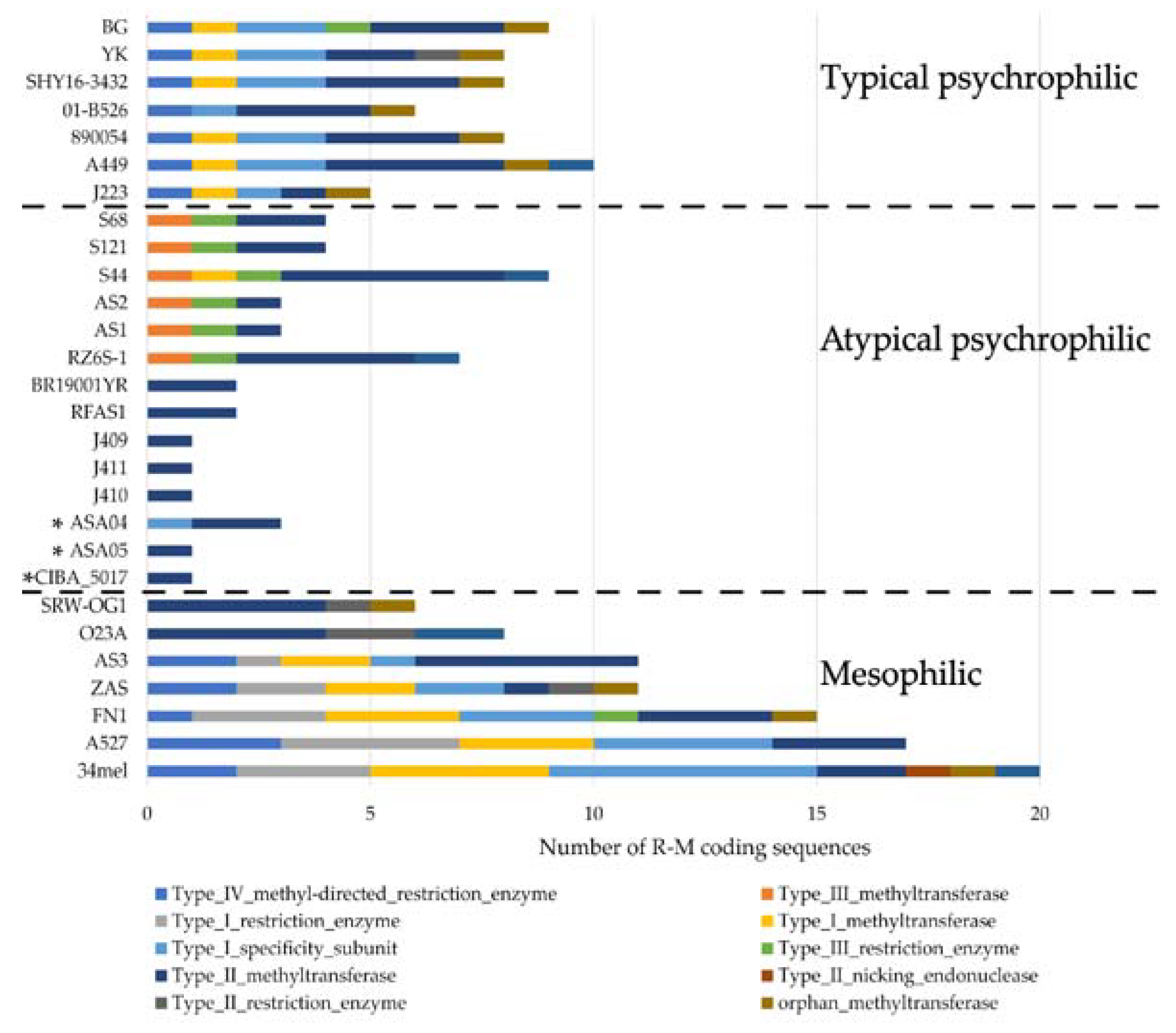
3.6. Insertion Sequences and Restriction-Modification System Abundance
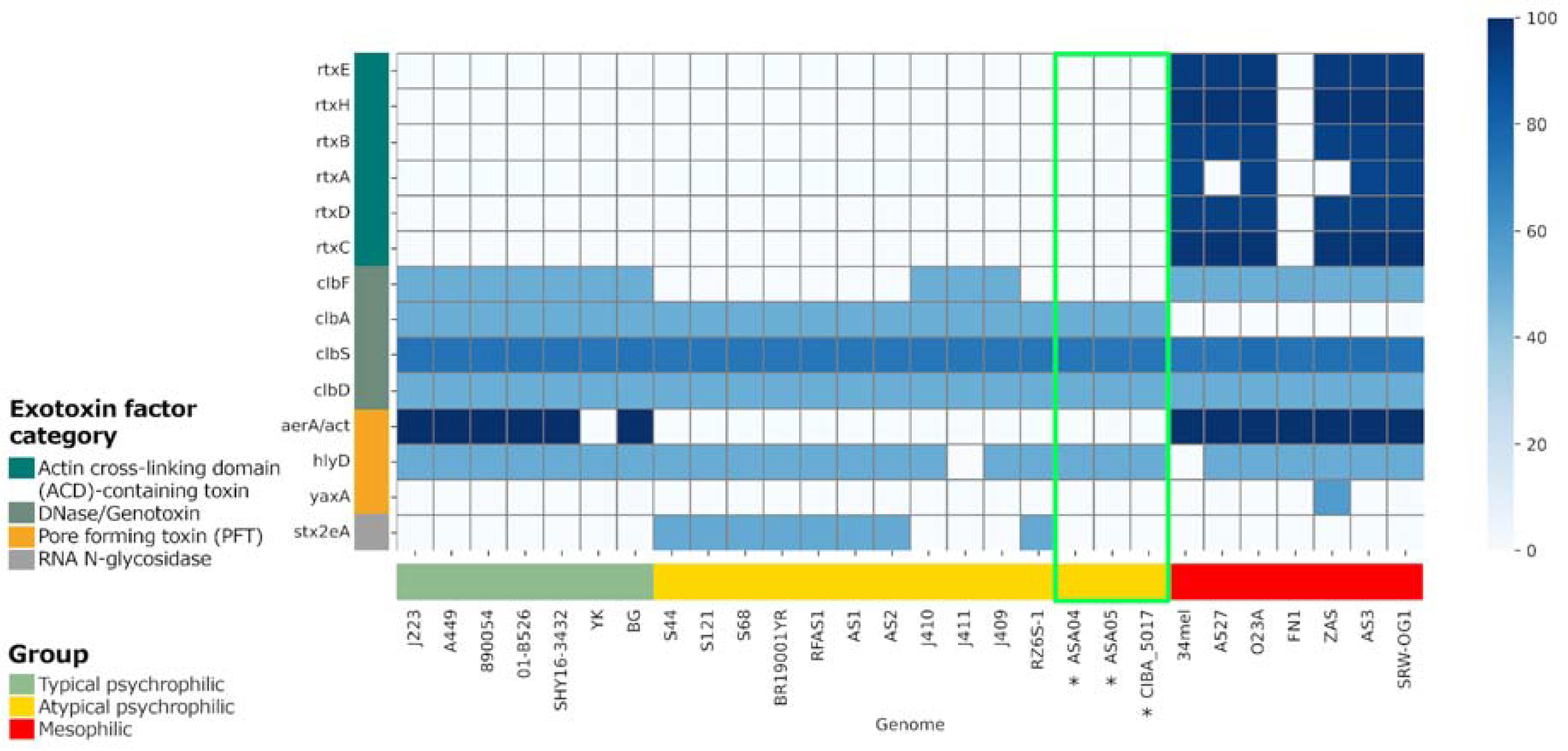
3.7. Virulence Factors
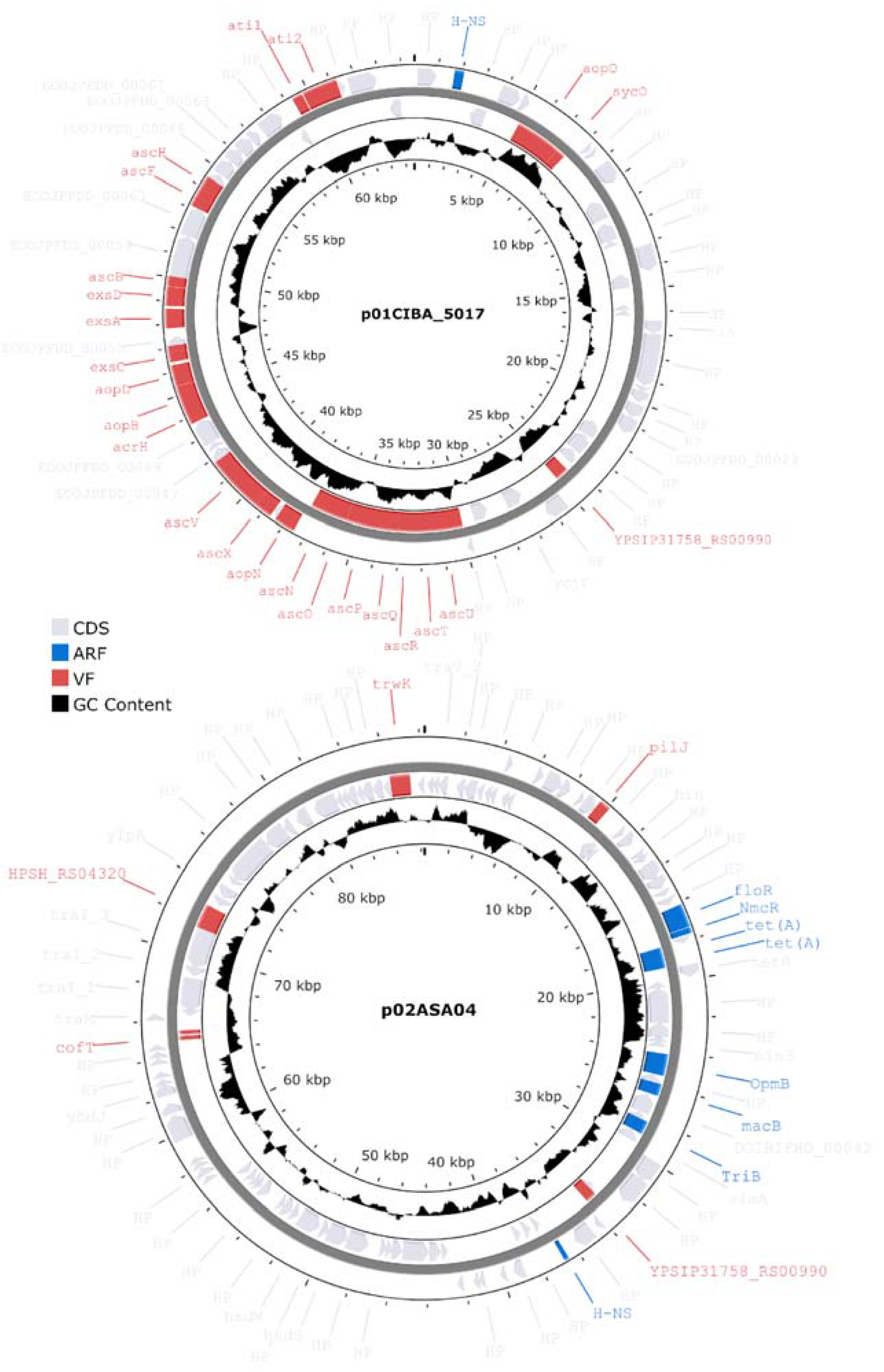
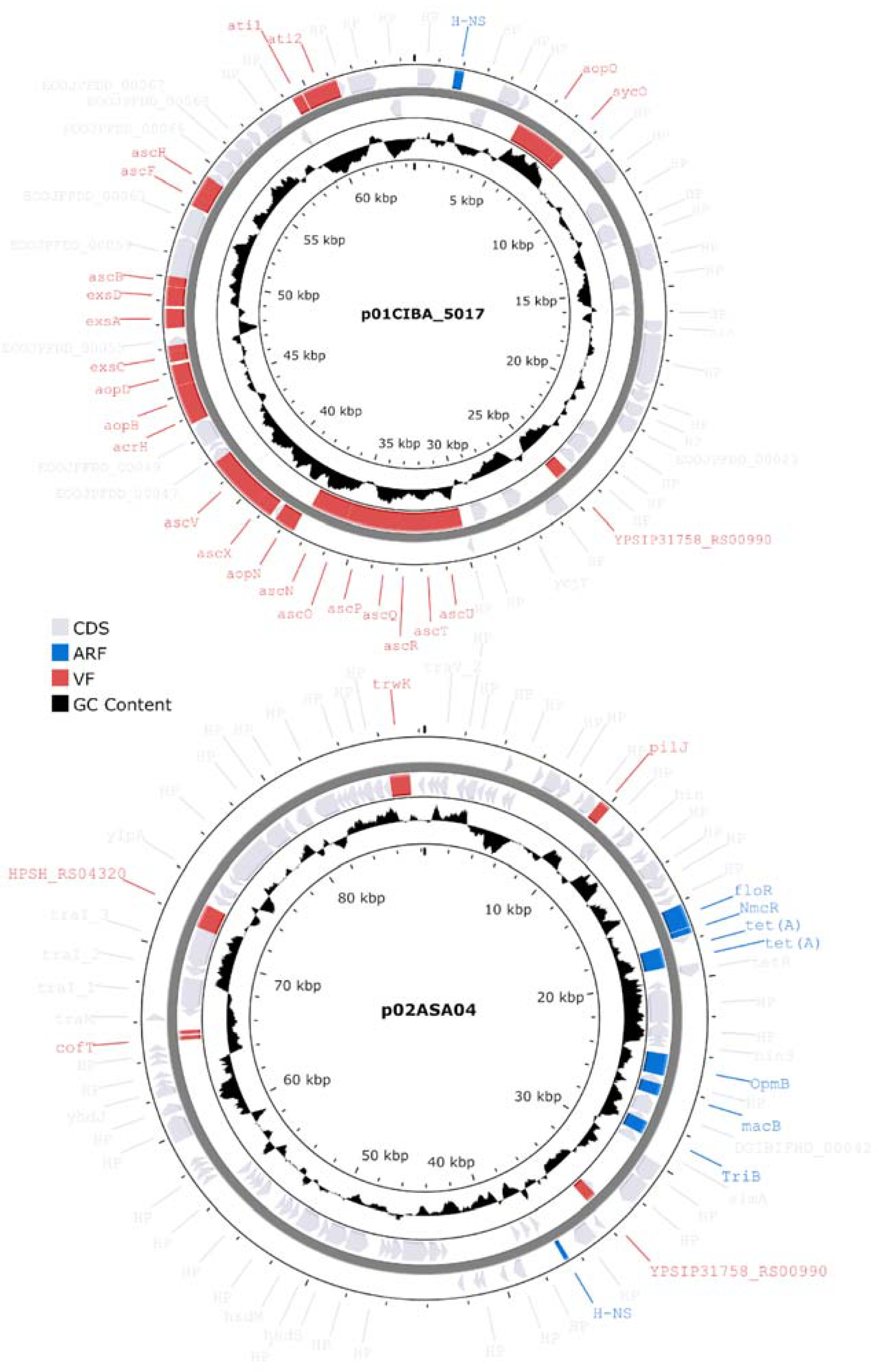
3.8. Characterization of Plasmids
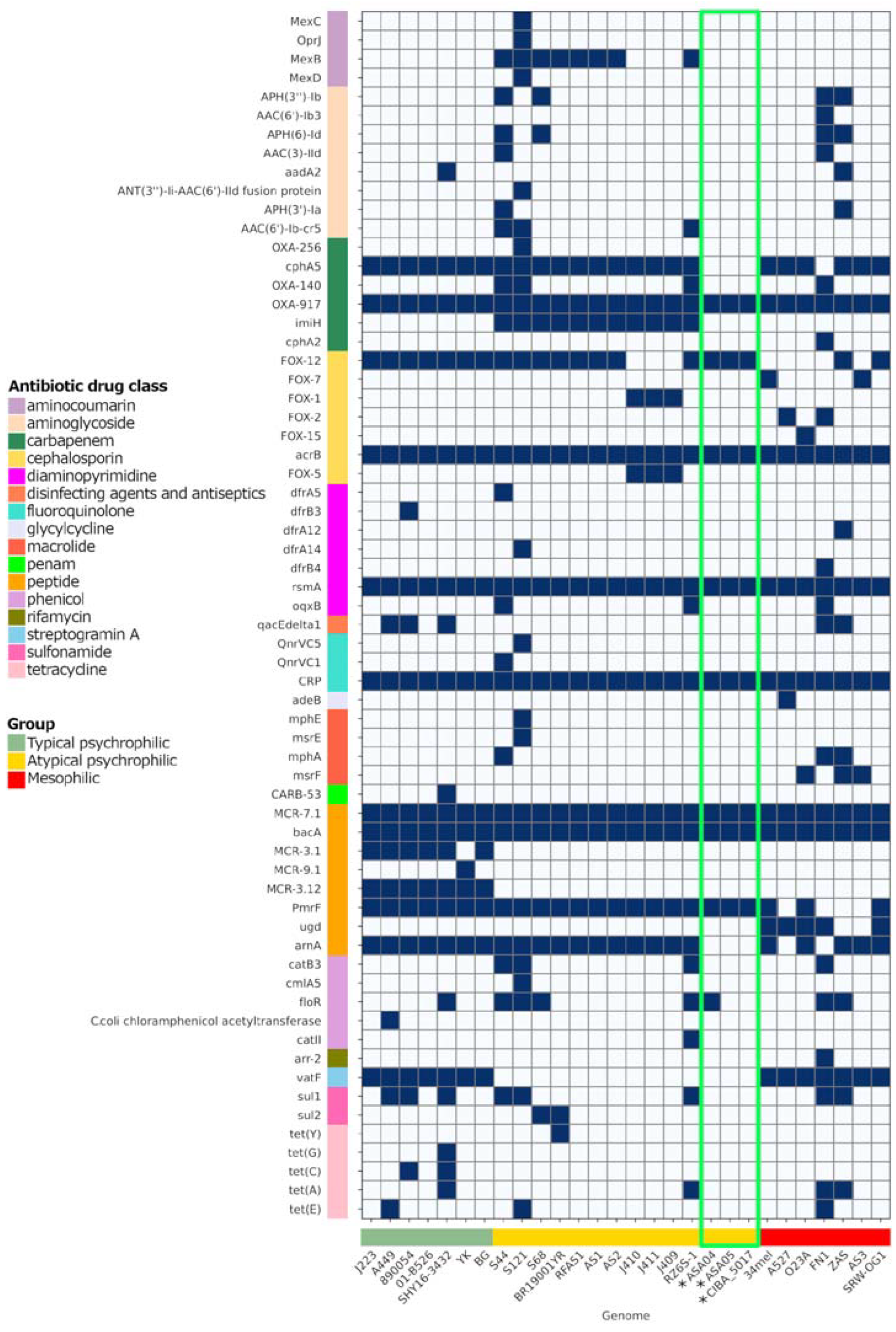
3.9. Antibiotic Resistance Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin-Carnahan, A.; Joseph, S.W. Aeromonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; Wiley: Hoboken, NJ, USA, 2015; pp. 1–44. ISBN 978-1-118-96060-8. [Google Scholar]
- Austin, D.A.; McIntosh, D.; Austin, B. Taxonomy of Fish Associated Aeromonas Spp., with the Description of Aeromonas salmonicida Subsp. Smithia Subsp. Nov. Syst. Appl. Microbiol. 1989, 11, 277–290. [Google Scholar] [CrossRef]
- Bernoth, E.-M. Furunculosis: Multidisciplinary Fish Disease Research; Academic Press: San Diego, CA, USA, 1997; ISBN 978-0-12-093040-1. [Google Scholar]
- Emmerich, R. Über Eine Durch Bacterien Erzeugte Seuche Unter Den Forellen. Arch. Hyg. Bakteriol. 1894, 21, 1–21. [Google Scholar]
- Kimura, T. A New Subspecies of Aeromonas salmonicida as an Etiological Agent of Furunculosis on “Sakuramasu” (Oncorhynchus masou) and Pink Salmon (O. Gorbuscha) Rearing for Maturity. Part 1. On the Morphological and Physiological Properties. Fish Pathol. 1969, 3, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T. A New Subspecies of Aeromonas salmonicida as an Etiological Agent of Furunculosis on “Sakuramasu” (Oncorhynchus masou) and Pink Salmon (O. Gorbuscha) Rearing for Maturity. Part 2. On the Serological Properties. Fish Pathol. 1969, 3, 45–52. [Google Scholar] [CrossRef]
- Mccarthy, D.H. Fish furunculosis caused by Aeromonas salmonicida var. Achromogenes. J. Wildl. Dis. 1975, 11, 489–493. [Google Scholar] [CrossRef]
- Smith, I.W. The Classification of “Bacterium salmonicida”. J. Gen. Microbiol. 1963, 33, 263–274. [Google Scholar] [CrossRef]
- Pavan, M.E.; Abbott, S.L.; Zorzópulos, J.; Janda, J.M. Aeromonas salmonicida Subsp. Pectinolytica Subsp. Nov., a New Pectinase-Positive Subspecies Isolated from a Heavily Polluted River. Int. J. Syst. Evol. Microbiol. 2000, 50, 1119–1124. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.A. Aeromonadaceae Representative (Aeromonas salmonicida). In Bacterial Fish Pathogens; Springer: Dordrecht, The Netherlands, 2012; pp. 147–228. ISBN 978-94-007-4883-5. [Google Scholar]
- Beaz, R.; Jos, M. Molecular Detection and Characterization of Furunculosis and Other Aeromonas Fish Infections. In Health and Environment in Aquaculture; Carvalho, E., Ed.; InTechOpen: London, UK, 2012; ISBN 978-953-51-0497-1. [Google Scholar]
- Menanteau-Ledouble, S.; Kumar, G.; Saleh, M.; El-Matbouli, M. Aeromonas Salmonicida: Updates on an Old Acquaintance. Dis. Aquat. Organ. 2016, 120, 49–68. [Google Scholar] [CrossRef]
- Vincent, A.T.; Rouleau, F.D.; Moineau, S.; Charette, S.J. Study of Mesophilic Aeromonas Salmonicida A527 Strain Sheds Light on the Species’ Lifestyles and Taxonomic Dilemma. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Gudmundsdottir, B.K.; Bjornsdottir, B. Aeromonas salmonicida and A. hydrophila. In Fish Viruses and Bacteria: Pathobiology and Protection; Woo, P.T.K., Cipriano, R.C., Eds.; CABI: Oxfordshire, UK, 2017; pp. 173–189. ISBN 978-1-78064-778-4. [Google Scholar]
- Cipriano, R.C.; Austin, B. Furunculosis and Other Aeromonad Diseases. In Fish Diseases and Disorders. Volume 3: Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; CABI: Oxfordshire, UK, 2011; pp. 424–483. ISBN 978-1-84593-554-2. [Google Scholar]
- Bravo, S. Atypical Furunculosis in Atlantic Salmon. Bull. Eur. Assoc. Fish Pathol. 2000, 20, 5. [Google Scholar]
- Godoy, M.; Gherardelli, V.; Heisinger, A.; Fernández, J.; Olmos, P.; Ovalle, L.; Ilardi, P.; Avendaño-Herrera, R. First Description of Atypical Furunculosis in Freshwater Farmed Atlantic Salmon, Salmo Salar L., in Chile. J. Fish Dis. 2010, 33, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Bravo, S.; Midtlyng, P.J. The Use of Fish Vaccines in the Chilean Salmon Industry 1999–2003. Aquaculture 2007, 270, 36–42. [Google Scholar] [CrossRef]
- SERNAPESCA. Informe Sanitario Con Informacion Sanitaria de Agua Dulce y Mar Año 2022; Departamento de Salud Animal, Subdireccion de Acuicultura, Servicio Nacional de Pesca y Acuicultura: Valparaíso, Chile, 2023. [Google Scholar]
- Löscher, T.; Prüfer-Krämer, L. Emerging and Re-Emerging Infectious Diseases. In Modern Infectious Disease Epidemiology; Krämer, A., Kretzschmar, M., Krickeberg, K., Eds.; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; pp. 39–67. ISBN 978-0-387-93834-9. [Google Scholar]
- Emond-Rheault, J.-G.; Vincent, A.T.; Trudel, M.V.; Brochu, F.; Boyle, B.; Tanaka, K.H.; Attéré, S.A.; Jubinville, É.; Loch, T.P.; Winters, A.D.; et al. Variants of a Genomic Island in Aeromonas salmonicida Subsp. Salmonicida Link Isolates with Their Geographical Origins. Vet. Microbiol. 2015, 175, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Emond-Rheault, J.-G.; Vincent, A.T.; Trudel, M.V.; Frey, J.; Frenette, M.; Charette, S.J. AsaGEI2b : A New Variant of a Genomic Island Identified in the Aeromonas salmonicida Subsp. Salmonicida JF3224 Strain Isolated from a Wild Fish in Switzerland. FEMS Microbiol. Lett. 2015, 362, fnv093. [Google Scholar] [CrossRef]
- Long, M.; Nielsen, T.K.; Leisner, J.J.; Hansen, L.H.; Shen, Z.X.; Zhang, Q.Q.; Li, A. Aeromonas salmonicida Subsp. Salmonicida Strains Isolated from Chinese Freshwater Fish Contain a Novel Genomic Island and Possible Regional-Specific Mobile Genetic Elements Profiles. FEMS Microbiol. Lett. 2016, 363, fnw190. [Google Scholar] [CrossRef]
- Vincent, A.T.; Intertaglia, L.; Loyer, V.; Paquet, V.E.; Adouane, É.; Martin, P.; Bérard, C.; Lami, R.; Charette, S.J. AsaGEI2d: A New Variant of a Genomic Island Identified in a Group of Aeromonas salmonicida Subsp. Salmonicida Isolated from France, Which Bears the pAsa7 Plasmid. FEMS Microbiol. Lett. 2021, 368, fnab021. [Google Scholar] [CrossRef]
- Gulla, S.; Bayliss, S.; Björnsdóttir, B.; Dalsgaard, I.; Haenen, O.; Jansson, E.; McCarthy, U.; Scholz, F.; Vercauteren, M.; Verner-Jeffreys, D.; et al. Biogeography of the Fish Pathogen Aeromonas salmonicida Inferred by vapA Genotyping. FEMS Microbiol. Lett. 2019, 366, fnz074. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Trudel, M.V.; Freschi, L.; Nagar, V.; Gagné-Thivierge, C.; Levesque, R.C.; Charette, S.J. Increasing Genomic Diversity and Evidence of Constrained Lifestyle Evolution Due to Insertion Sequences in Aeromonas salmonicida. BMC Genom. 2016, 17, 44. [Google Scholar] [CrossRef]
- Vasquez, I.; Hossain, A.; Gnanagobal, H.; Valderrama, K.; Campbell, B.; Ness, M.; Charette, S.J.; Gamperl, A.K.; Cipriano, R.; Segovia, C.; et al. Comparative Genomics of Typical and Atypical Aeromonas salmonicida Complete Genomes Revealed New Insights into Pathogenesis Evolution. Microorganisms 2022, 10, 189. [Google Scholar] [CrossRef]
- Long, M.; Fan, H.; Gan, Z.; Jiang, Z.; Tang, S.; Xia, H.; Lu, Y. Comparative Genomic Analysis Provides Insights into Taxonomy and Temperature Adaption of Aeromonas salmonicida. J. Fish Dis. 2023, 46, 545–561. [Google Scholar] [CrossRef]
- Roh, H.; Kannimuthu, D. Comparative Resistome Analysis of Aeromonas Species in Aquaculture Reveals Antibiotic Resistance Patterns and Phylogeographic Distribution. Environ. Res. 2023, 239, 117273. [Google Scholar] [CrossRef] [PubMed]
- Bickle, T.A.; Krüger, D.H. Biology of DNA Restriction. Microbiol. Rev. 1993, 57, 434–450. [Google Scholar] [CrossRef] [PubMed]
- Pingoud, A.; Fuxreiter, M.; Pingoud, V.; Wende, W. Type II Restriction Endonucleases: Structure and Mechanism. Cell. Mol. Life Sci. 2005, 62, 685–707. [Google Scholar] [CrossRef]
- Roberts, R.J. A Nomenclature for Restriction Enzymes, DNA Methyltransferases, Homing Endonucleases and Their Genes. Nucleic Acids Res. 2003, 31, 1805–1812. [Google Scholar] [CrossRef] [PubMed]
- Murray, N.E. Immigration Control of DNA in Bacteria: Self versus Non-Self. Microbiology 2002, 148, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Valdes, N.; Espinoza, C.; Sanhueza, L.; Gonzalez, A.; Corsini, G.; Tello, M. Draft Genome Sequence of the Chilean Isolate Aeromonas Salmonicida Strain CBA100. FEMS Microbiol. Lett. 2015, 362, fnu062. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Noga, E.J. Fish Disease: Diagnosis and Treatment, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2010; ISBN 978-0-8138-0697-6. [Google Scholar]
- Prophet, E.; Armed Forces Institute of Pathology (U.S.). Laboratory Methods in Histotechnology, 7th ed.; American Registry of Pathology: Arlington, VA, USA, 1992. [Google Scholar]
- Keeling, S.E.; Brosnahan, C.L.; Johnston, C.; Wallis, R.; Gudkovs, N.; McDonald, W.L. Development and Validation of a Real-time PCR Assay for the Detection of Aeromonas Salmonicida. J. Fish Dis. 2013, 36, 495–503. [Google Scholar] [CrossRef]
- Zhao, J.; Vendramin, N.; Cuenca, A.; Polinski, M.; Hawley, L.M.; Garver, K.A. Pan-Piscine Orthoreovirus (PRV) Detection Using Reverse Transcription Quantitative PCR. Pathogens 2021, 10, 1548. [Google Scholar] [CrossRef]
- Snow, M.; McKay, P.; McBeath, A.J.A.; Black, J.; Doig, F.; Kerr, R.; Cunningham, C.O.; Nylund, A.; Devold, M. Development, Application and Validation of a Taqman Real-Time RT-PCR Assay for the Detection of Infectious Salmon Anaemia Virus (ISAV) in Atlantic Salmon (Salmo salar). Dev. Biol. 2006, 126, 133–145, discussion 325–326. [Google Scholar]
- Ingerslev, H.-C.; Rønneseth, A.; Pettersen, E.F.; Wergeland, H.I. Differential Expression of Immune Genes in Atlantic Salmon (Salmo Salar L.) Challenged Intraperitoneally or by Cohabitation with IPNV. Scand. J. Immunol. 2009, 69, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Austin, B. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2016; ISBN 978-3-319-32673-3. [Google Scholar]
- Paterson, W.D.; Douey, D.; Desautels, D. Isolation and Identification of an Atypical Aeromonas salmonicida Strain Causing Epizootic Losses Among Atlantic Salmon (Salmo salar) Reared in a Nova Scotian Hatchery. Can. J. Fish. Aquat. Sci. 1980, 37, 2236–2241. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and Accurate Long-Read Assembly via Adaptive k -Mer Weighting and Repeat Separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLOS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Bayliss, S.C.; Thorpe, H.A.; Coyle, N.M.; Sheppard, S.K.; Feil, E.J. PIRATE: A Fast and Scalable Pangenomics Toolbox for Clustering Diverged Orthologues in Bacteria. GigaScience 2019, 8, giz119. [Google Scholar] [CrossRef]
- Seemann, T. Snippy: Rapid Haploid Variant Calling and Core Genome Alignment. 2015. Available online: https://github.com/tseemann/snippy(accessed on 20 October 2023).
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A Comparative Pathogenomic Platform with an Interactive Web Interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2019, 48, gkz935. [Google Scholar] [CrossRef]
- Siguier, P. ISfinder: The Reference Centre for Bacterial Insertion Sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. REBASE: A Database for DNA Restriction and Modification: Enzymes, Genes and Genomes. Nucleic Acids Res. 2023, 51, D629–D630. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-Depth Characterization and Visualization of Bacterial Genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed]
- The Pandas Development Team. Pandas-Dev/Pandas: Pandas. 2023. Available online: https://zenodo.org/doi/10.5281/zenodo.3509134 (accessed on 2 October 2023).
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Waskom, M. Seaborn: Statistical Data Visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Roder, T. Flower-Plot. 2021. Available online: https://github.com/MrTomRod/flower-plot (accessed on 20 October 2023).
- Bruno, D.W.; Noguera, P.A.; Poppe, T.T. A Colour Atlas of Salmonid Diseases; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-2009-1. [Google Scholar]
- Sibley, M.H. Cassette-like Variation of Restriction Enzyme Genes in Escherichia coli C and Relatives. Nucleic Acids Res. 2004, 32, 522–534. [Google Scholar] [CrossRef]
- Corvaglia, A.R.; François, P.; Hernandez, D.; Perron, K.; Linder, P.; Schrenzel, J. A Type III-like Restriction Endonuclease Functions as a Major Barrier to Horizontal Gene Transfer in Clinical Staphylococcus Aureus Strains. Proc. Natl. Acad. Sci. USA 2010, 107, 11954–11958. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case ID | 4978 | 4988 | 5131 | 7003 | 7031 | 7081 | 7088 | 7124 | 7167 | 7175 | 7174 | 7215 | 7216 | 7214 |
| Date | 10-Mar | 14-Mar | 11-May | 23-May | 03-Jun | 23-Jun | 30-Jun | 21-Jul | 08-Aug | 12-Aug | 12-Aug | 18-Aug | 24-Aug | 25-Aug |
| Region | X | X | X | X | X | XII | XII | XII | X | X | X | XII | XII | X |
| Salinity | FW | FW | BW | FW | FW | BW | BW | BW | BW | BW | FW | BW | BW | BW |
| Development stage | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt |
| Fish sampled | 20 | 7 | 10 | 30 | 6 | 7 | 7 | 10 | 10 | 11 | 24 | 1 | 1 | 1 |
| Average ASA Ct | 22.8 | 26.1 | 29.2 | 28.4 | 23.8 | 27.5 | 26.2 | 24.4 | 27.1 | 29.8 | 21.4 | 26.1 | 32.8 | 22.5 |
| Coinfection | PRV | IPNV | IPNV | R. salm | PRV | IPNV-F.psy | IPNV-PRV | IPNV-PRV | - | - | IPNV-PRV | - | - | - |
| Bacterial isolation | Positive | Positive | Positive | Negative | Negative | Negative | Negative | Negative | Positive | Negative | Negative | Negative | Negative | Negative |
| GenBank accession | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Case ID | 7198 | 7213 | 7217 | 7385 | 7388 | 7387 | 7412 | 7416 | 7435 | CIBA-5017 | ASA04 | ASA05 | ||
| Date | 26-Aug | 26-Aug | 06-Sept | 20-Oct | 21-Oct | 21-Oct | 27-Oct | 28-Oct | 04-Nov | 25-Mar | 22-Nov | 23-Nov | ||
| Region | X | X | X | X | X | X | X | X | X | X | X | X | ||
| Salinity | FW | FW | FW | FW | BW | FW | FW | FW | BW | FW | FW | FW | ||
| Development stage | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Smolt | Smolt | Smolt | ||
| Fish sampled | 10 | 1 | 7 | 24 | 24 | 26 | 15 | 10 | 15 | 20 | 25 | 11 | ||
| Average ASA Ct | 29.6 | 26.2 | 23.2 | 30.6 | 26.6 | 24.3 | 30.8 | 29.1 | 27.0 | 28.8 | 29.7 | 26.1 | ||
| Coinfection | IPNV | - | IPNV | - | - | - | - | PRV | PRV | - | IPNV-PRV | IPNV-PRV | ||
| Bacterial isolation | Positive | Negative | Negative | Negative | Negative | Positive | Negative | Negative | Negative | Positive | Positive | Positive | ||
| GenBank accession | - | - | - | - | - | - | - | - | - | CP139910 | CP139915 | CP139912 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godoy, M.; Montes de Oca, M.; Suarez, R.; Martinez, A.; Pontigo, J.P.; Caro, D.; Kusch, K.; Coca, Y.; Bohle, H.; Bayliss, S.; et al. Genomics of Re-Emergent Aeromonas salmonicida in Atlantic Salmon Outbreaks. Microorganisms 2024, 12, 64. https://doi.org/10.3390/microorganisms12010064
Godoy M, Montes de Oca M, Suarez R, Martinez A, Pontigo JP, Caro D, Kusch K, Coca Y, Bohle H, Bayliss S, et al. Genomics of Re-Emergent Aeromonas salmonicida in Atlantic Salmon Outbreaks. Microorganisms. 2024; 12(1):64. https://doi.org/10.3390/microorganisms12010064
Chicago/Turabian StyleGodoy, Marcos, Marco Montes de Oca, Rudy Suarez, Alexis Martinez, Juan Pablo Pontigo, Diego Caro, Karina Kusch, Yoandy Coca, Harry Bohle, Sion Bayliss, and et al. 2024. "Genomics of Re-Emergent Aeromonas salmonicida in Atlantic Salmon Outbreaks" Microorganisms 12, no. 1: 64. https://doi.org/10.3390/microorganisms12010064
APA StyleGodoy, M., Montes de Oca, M., Suarez, R., Martinez, A., Pontigo, J. P., Caro, D., Kusch, K., Coca, Y., Bohle, H., Bayliss, S., Kibenge, M., & Kibenge, F. (2024). Genomics of Re-Emergent Aeromonas salmonicida in Atlantic Salmon Outbreaks. Microorganisms, 12(1), 64. https://doi.org/10.3390/microorganisms12010064